Inleiding
Ecologische observatie’s, literatuurstudie en een eerste kweekverslag rond Ambystoma macrodactylum macrodactylum & A. m. columbianum uit Oregon, USA.

Ambystoma macrodactylum behoort tot de familie van de molsalamanders ( Ambystomatidae). Deze familie omvat een flink aantal soorten die bijna allemaal opvallen door hun mooie kleur en terzelfdertijd hun vrij verscholen levenswijze. Merendeel van deze soorten leven tijdens de landfase ondergronds en komen in het voorjaar tevoorschijn om zich dan vrij massaal voort te planten in het water.
De larven zijn in de poelen en plassen waar geen vissen voorkomen ook vaak de toppredator van de voedselketen. De naam macrodactylum ( macro = groot, dactylus = ) dankt het dier aan zijn verhoudingsgewijze grote tenen. Ook in het Engels wordt hij de ‘Longtoed’ salamander genoemd. Een echte nederlandse naam bestaat vermoedelijk nog niet, mede omdat deze dieren tot op heden eigenlijk vrij zelden beschikbaar waren. Hopelijk komt door de reeds herhaalde kweekresultaten een verandering in.
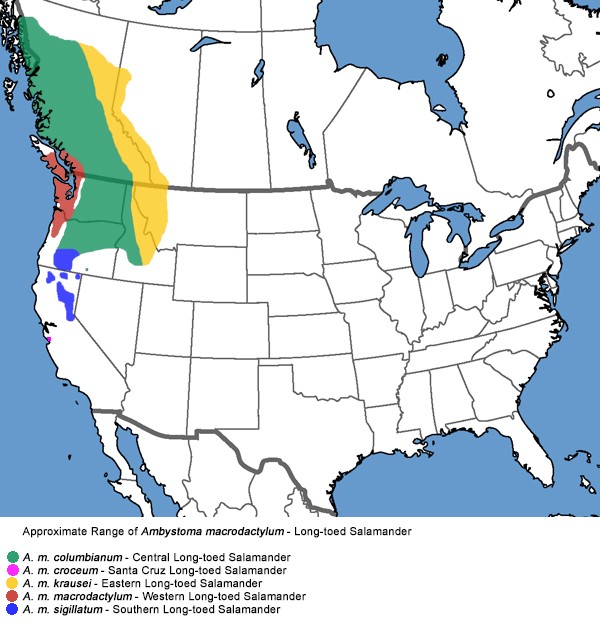
Ondersoorten
1) Ambystoma macrodactylum macrodactylum : De felgele streep van de jonge dieren wordt doffer in volwassen dieren. Mijn nakweek vertoonde enkel een felgele tekening in het eerste levensjaar. Naar de randen toe lost de streep op in de grondkleur, waardoor een ‘echte’ overgang tussen de beide kleuren moeilijk te ontwaren is. Het midden van deze streep breekt met het ouder worden als het ware open en de grondkleur vorm dan een soort zwarte middellijn die typisch is voor deze vorm. De staart bezit geen verkleuring langsheen de zijkanten.

2) Ambystoma macrodactylum sigillatum : De kleur van de band is in deze ondersoort het felst van alle ondersoorten. De vorm van de streep is vrij variabel en doorgaans smal en kan zowel bestaan uit een diep ingesneden rechte lijn als uit een verzameling van puntjes en vlekken. De vlekken op de snuit zijn meestal kleiner dan deze achter de ogen. De buikpartij is eerder roze-achtig en doorschijnend. Men kan er de ingewanden doorheen zien.

3) Ambystoma macrodactylum columbianum : De felgele lijn is ononderbroken en vertoont geen insnijdingen, in tegenstelling met de nominaatvorm hebben we geen donkere middenlijn. Op de kop (tussen de neusgaten) verbreedt de gele ruglijn. Zowel op de kop, snuit en oogleden vertonen vele vlekken. Typisch vindt is de sterke gele kleur op de bovenzijde van de staart en soms zelfs‘overloopt’ op de zijkanten ervan.

4) Ambystoma macrodactylum croceum : Deze ondersoort werd pas in 1956 door Russell en Anderson ontdekt in Santa Cruz, Sierra Nevada. Dit kustelijk verspreidingsgebied ligt zowat 600 km ten Zuiden van de andere kustelijke ondersoorten en zo’n 250 km ten Zuidwesten van de andere Sierra Nevada populatie’s., waarvan het geïsoleerd is door de Grote Centrale vallei van California. Croceum dieren bezitten achter de ogen typische oranje vlekken. De ononderbroken rugstreep is bleekoranje en vertoont de neiging om op de kop opgebroken te worden in kleinere verspreide vlekjes, die meermaals ontbreken net achter de ogen.
Deze bijzonder fraaie ondersoort is echter met uitsterven bedreigd en geniet dan ook de meer dan nodige bescherming. Door deze status en het toch wel bijzonder eigenaardige verspreidingsgebied werd behoorlijk wat onderzoek gedaan oa. op het vlak van voortplanting & voedsel. Hierdoor is over deze zeldzame ondersoort eigenlijk meer bekend dan over zijn meer algemene neven. Deze dieren zijn uiteraard voor de liefhebber niet beschikbaar.

5) Ambystoma macrodactylum krausei : Krausei bezit een rechte analine-gele ononderbroken ruglijn die doorloopt van de snuit tot het staarteinde . Deze band is vrij dun en behoorlijk rechtlijnig. In tegenstelling met andere ondersoorten wordt deze op de snuit niet opgebroken in kleinere vlekjes, wel wordt ze met name in de omgeving van de ogen behoorlijk breder. De vlekken op de oogleden zijn geïsoleerd en zijn hierdoor sterk geaccentueerd. Met een beetje verbeelding denk je aan de uitklapbare koplampen v/e sportwagen.
De eenvoud van deze rechtlijnige indeling wordt echter deels teniet gedaan doordat in de overlappende gebieden tussenvormen voorkomen. In Olympic Peninsula vinden we bvb. exemplaren van de nominaatvorm die eenzelfde rugstreep vertonen als A. macrodactylum columbianum. In Klamath en Lake Counties ( Zuid Centraal Oregon) vinden we dan weer tussenvormen van A. sigillatum en A. columbianum.

Bemerkingen aan het biotoop van macrodactylum Columbianum in de Columbia River Gorge.
Tijdens de maand april ’98 kon ik met de medehulp van Kurtz Russell, observaties verrichten in Oregon. In de Columbia River Gorge, ontdekten we tussen de ontelbare Hyla regilla larven een groot aantal nog kleine Ambystoma macrodactylum columbianum larven (1,8 tot 2 cm) in een niet-permanente waterplas. De lengte Deze was gelegen in volle zon langsheen de rijweg. De vegetatie bestond vnlk. uit een slijmerig wilde algengroei tussen een aantal grasachtige moerasplanten. We noteerden geen echte waterplanten, vermoedelijk omdat de plas in de zomer uitdroogt.
De watertemperatuur bedroeg 19°C, de luchttemperatuur ongeveer 30°C. Volwassen dieren ontbraken volledig, deze hadden zich bij deze temperaturen –die niet zo veel onder de kritische temperatuur lagen- waarschijnlijk ondergronds teruggetrokken in de aangrenzende grasvelden. Hiervoor situeren ze zich in de eerste 40 cm diepte en gebruiken frequent tunnels van kleine knaagdieren, waarvan ze wegens hun huidsecreet niets hoeven te vrezen..
Anderson J.D. voerde hierrond overigens heel wat onderzoeken uit ao. de predatie van Ambystoma macrodactylum croceum door de mol (Scapanus latimanus). Deze anders erg vraatzuchtige mol, die zonder schralen in zijn dieet ook wormachtige Batrachoseps salamanders opneemt, weigerde juveniele en zelfs op een pincet aangeboden larven van A. croceum. Onder heen enkel beding raakte de uitgehongerde mol de salamanders aan, beter nog, hij kroop zelfs terug indien de buit hem te dicht werd aangeboden.
Bemerkingen aan het biotoop van A. macrodactylum macrodactylum in Eugene, Oregon
In diezelfde periode vonden we in een open grasveld Eugene een klein grachtje van het permanente type. De afwezigheid van waterplanten verklaarde Russell was te ‘danken’ aan een opkuisbeurt in het vorige jaar. Het water was licht troebel, men kon enkel grote vormen waarnemen. Op sommige plaatsen begon het kransvederkruid (Myriophylum sp.) zich op een bescheiden manier terug te ontwikkelen. Langsheen de zijkanten tussen de grassen vonden we kleine klompjes eitjes die zowel van A. macrodactylum macrodactylum als Hyla regilla konden zijn. Beide dieren zettten nl. op dergelijke wijze hun zachte gelei-achtige eitjes af.
Hoogstwaarschijnlijk betrof het echter Hyla regilla aangezien Ambystoma macrodactylum macrodactylum eigenlijk veel vroeger afzet. Na een paar schepbeurten die vooral onder de weinige waterplanten vondsten opleverden, waren we een 20 tal Ambystoma macrodactylum macrodactylum larven rijker. De dieren maten 4.5 tot 5 cm lang en waren hiermee dichtbij hun metamorfose belandt. Net zoals bij het Ambystoma macrodactylum columbianum biotoop noteerden we de massale aanwezigheid van Hyla regilla larfjes. Gezien de snelle ontwikkeling van de Ambystoma macrodactylum macrodactylum larven lijkt het mogelijk dat deze kikkervisjes of zelfs de eitjes een veelgebruikte voedselbron zijn …
Langsheen de zijkanten van een moeras vonden we een 10 tal even ongelukkige Tamnophis sirtalis , ze hadden het ook graag anders gezien, zij het om duidelijk minder vredelievende redenen .
Een grote afvalhoop achter deze gracht was nog een stille getuige van de verbouwingswerken die vorige jaren aan de nabije kerk waren uitgevoerd. Deze opeengestapelde afval was wild overgroeid met een ondoordringbare jungle van (klim)planten. Hoogstwaarschijnlijk keken enkele adulte dieren vanuit hun veilige schuilplaats onze verwoede maar ijdele pogingen met veel plezier na.
We bekeken nog een aantal moerasgebieden, tijdelijke plassen en poelen, maar in al deze gebruikelijke biotopen, die volgens Russel in het vroege voorjaar wel A. macrodactylum verbergen, kwamen we echter te laat. De lente van ’98 was ongebruikelijk warm geweest, terwijl de temperaturen normaal 12 tot 15°C waren, wezen sommige metingen tot wel 32°C aan. Onder dergelijke temperaturen waren de larven vermoedelijk al gemetamorfoseerd en de adulte dieren hadden hun naam molsalamander terug alle eer aangedaan.
Het is alsof Taricha granulosa & A. macrodactylum, die beide hun eitjes afzetten in stilstaand water, elkaar mijden. In geen enkel van de zowat 15 door ons onderzochte poelen vonden we de larven van beide soorten samen. Terwijl Taricha granulosa blijkbaar eerder diepere & permanente waterplassen gebruikt, lijkt Ambystoma macrodactylum macrodactylum zich hoofdzakelijk te beperken tot die waterplassen die tijdens de zomer wel uitdrogen … In een diepere gracht van het permanente type vonden we bvb. in Eugene tot 5,5cm grote Taricha granulosa larven.
Inrichting terrarium
A. macrodactylum heeft een vrij hoge vochtnood, die kan wel verschillen per ondersoort. Tijdens een onderzoek naar de vochtigheids behoefte van 3 ondersoorten door Anderson (1972) blijkt dat A. croceum (die tevens in een warmer klimaat vertoeft) een behoorlijk lagere vochtnood bezit dan de nominaatvorm en A. krausei. Deze vochtnood verschilt ook per periode : terug bij A. croceum lag deze tijdens de voorplantingsfase, heel wat hoger dan erbuiten.
Uitgaande van mijn terrarium ervaringen met en de observatie’s aan hun leefomgeving hou ik de dieren in een soort van waterkant-biotoop. De bodem van de bak bestaat uit (vijver)grind wat naar achter toe opgehoogd is. Op deze verhoging – die ongeveer de helft van de bak beslaat- worden brokstukken van snelbouwstenen gelegd tot een paar centimeter van de rand. Deze uit lavasteen gemaakte stenen hebben grote open ruimtes waarin de dieren zich graag verschuilen.
Met op elkaar geplaatste dakpannen kan vermoedelijk ook hetzelfde effect verkregen worden. Tussen en op deze stenen wordt vervolgens spaghnumachtig mos gelegd, wat ik tijdens de lente en herfst tussen de beschaduwde plaatsen van het gazon uithaal. De stenen staan met hun voeten in het water en leveren door hun capillaire werking de nodige vochtigheid af aan het mos. De bak wordt volledig afgesloten met een dekglas en in een ietwat beschaduwde hoek geplaatst.
Op deze wijze wordt een vochtig omgeving met een vrij stabiele temperatuur gecreëerd. Enkele varens en indien voldoende licht aanwezig is, wat Ficus repens geven een dergelijke bak een natuurlijk uitzicht. Er dient met zorg op gezien te worden dat het directe zonlicht de bak niet raakt, anders stijgen de temperaturen (te) snel en dat kunnen deze dieren nu net niet hebben. Howard & Wallace (1983) bepaalden proefondervindelijk dat de kritische maximale temperatuur (d.i. de temperatuur waarbij de dieren afsterven) tussen de 33.2°C en 35 °C ligt, met een kleine variatie, afhankelijk van de hoogte van de vindplaats en de oorspronkelijke temperatuur waarbij de dieren initieel gehouden werden.
Voor het correct houden van deze soort dienen dergelijke hoge of snelle opsprongen in temperatuur (vnlk in de zomer) met zorg vermeden te worden. Dit kan gedeeltelijk opgevangen worden door het inbouwen van ondergrondse schuilplaatsen, wat dan ook een benadering is van hun natuurlijke levenswijze. Na enkele minder geslaagde try-outs, hou ik mijn dieren nu al enkele jaren succesvol op deze wijze, zonder uitval. Bij andere opstellingen oa. een bosbiotoop liep al vrij snel iets mis. De dieren schijnen net als mijn Hynobius een dergelijke drijfnatte omgeving erg te appreciëren. Ze zijn wel vrij gevoelig voor hogere temperaturen (> 25°C) en dienen derhalve best bij ‘veilige’ lagere temperaturen gehouden te worden
Voedsel
Anderson (1968) bestudeerde het voedingsgewoontes van volwassen en larven van 2 A. macrodactylum – ondersoorten, hieruit blijkt dat Arthropoden mn. Isopoden 50% van het voedsel uitmaken voor A. croceum. Vooral in de drogere periode is dit dier aangewezen op deze dieren, die dan in dezelfde vochtige omgeving in grote getallen voorkomen. In de nattere periodes worden duizendpoten, kevertjes, spinnen en wormen verorberd. Het A. siggilatum dieet bestaat daarentegen vnlk uit spinnen en insecten ipv isopoden.
In gevangenschap voer ik mijn volwassen dieren vnlk regenwormen, naaktslakken, meelwormpjes, muggenlarven, buffalo wormen, pissebedden, duizendpoten en vliegenmaden. Het zijn vraatzuchtige rover en de prooien worden tevens fel achterna gejaagd. In de plassen waar vissen afwezig zijn vormen de larven de toppredator. De juveniele dieren bvb. kijken telkens weer met veel interesse omhoog als ik het deksel van de bak oplicht. Tijdens het voederen komen deze dan zonder schroom uit hun schuilplaats om zich –letterlijk- op de prooidieren te storten. Terwijl het normaal vrij verdraagzame dieren zijn t.o.v. elkaar, bijten ze tijdens het voederen wild om zich heen om zo een hapje voor zichzelf te kunnen ‘reserveren’.
Voortplanting
In de natuur is Ambystoma macrodactylum een van de eerste actieve amfibieën. Men vindt ze reeds in plassen die nog overdekt zijn met ijs. Het zal dan ook niet verwonderlijk zijn dat deze dieren ook in gevangenschap reeds bij de prille eerste lentezon ‘in hun vel’ voelen … Je vindt de anders zo schuchtere dieren plots samen in het water. Eerst komen de mannetjes, daarna pas de vrij dikke zwangere vrouwtjes … net zoals in de vrije natuur.

De buikpartij van de dieren is quasi transparant zodat het controleren op eitjes bij de vrouwtjes een makkelijke klus is : je ziet de witte ronde wolken dwars doorheen de huid.Tijdens de herfst ’95 werden de dieren binnengebracht in een onverwarmde (huis)kamer. De watertemperaturen schommelden tussen 12 en 14°C. Na raadpleging van de literatuur ( K. Rimmp) bleken de dieren tot voortplanting gebracht te kunnen worden door een plotse koude vochtige lucht. Aangezien ze volgens diverse naslagwerken de kou goed kunnen verdragen hoefde ik ook niet bang te zijn om ze onder te brengen ik het tuinhuis, wat uiteindelijk op 14/1/96 gebeurde. In een aquarium van 80 op 35 cm plaatste ik 3 duidelijk zwangere vrouwtjes met ongeveer een 10 tal bronstige mannetjes.
De temperaturen bedroegen toen ongeveer 4°C. Door de geringe hoogte van de bak (ongeveer 20 cm) en het afsluiten ervan met een glasplaat bekwam ik de koele en vochtige (ijs)koude lucht. De dieren verbleven overdag op het land tussen de stenen. Slechts ‘s nachts gingen ze te water. Op 20 januari ontdekte ik reeds het eerste legsel van 94 eieren, die bijna allemaal werden afgezet op de drijfplanten (zie dia’s), meerbepaald op de wortels en langsheen de stengels werden hier en daar reeksen eieren gekleefd. Bij afwezigheid van planten in de bak denk ik dat een stevig lint of koord ook gebruikt kan worden. In Amerika gebruikt men deze methode nogal, omdat je dan gemakkelijk de eitjes kunt verwijderen door langsheen het lint met je vingers te glijden. Vooral bij A. talpoideum wordt deze truc gebruikt.
Ook op de overhangende Ficus repens vond ik (enkele) eitjes. Deze zijn vrij klein en tweekleurig. De bovenzijde is zwartbruin, de onderzijde is gebroken wit. Misschien speelt deze tweekleurigheid een rol in de intensiteit van het ontvangen licht voor de kern. Bovenaan wordt veel licht gefilterd door de donkere kleur, onderaan laat de lichte kleur meer licht door. Als resultaat krijgt de kern dan een evenredige lichtinval. Of hypothetiseer ik er maar op los ? Het viel me ook op dat de eitjes vrij dicht tegen het wateroppervlak werden afgezet. De manier van eiafzet kwam goed overeen met doe van bvb. de Axolotl (Ambystoma mexicanum) een mexicaanse grotere neef.
Op 22 januari werden ongeveer 31 eitjes in de onverwarmde kamer binnengebracht. De watertemperatuur was op dat moment 15°C. Er werden nog 64 eitjes in de afzetbak achtergelaten. Ik kon geen vraatzucht bij de ouderdieren waarnemen, ze lieten de eieren ongemoeid.
Op 27 januari vertoonden de eitjes reeds een kommavormige ontwikkeling. De watertemperatuur was toen 12°C. De achtergebleven eitjes in het tuinhuis vertoonden bij 2 tot 4°C nog geen enkel teken van evolutie. Er werden uiteindelijk geen verdere eitjes meer afgezet. Mogelijke oorzaken : de sterke daling van de buitentemperatuur (van 6 tot –6°c overdag), ofwel de te lage waterstand in de bak (6cm) ; wegens tijdsgebrek had ik nog geen kans gezien om het water wat gebruikt werd voor de verwijdering van de 31 eitjes weer aan te vullen. Aangezien de eitjes binnenshuis probleemloos en tegen grote snelheid evolueerden werden de overige eitjes ook binnengebracht op 29 januari.
Losgekomen eitjes die op de grond van de bak vallen leken minstens achterop te lopen in evolutie, het merendeel hiervan stierf af. Grote licht en/of zuurstofbehoefte ?
Opkweek : larven
Het opkweken van de larven bleek vrij eenvoudig. Daar in gevangenschap de concentratie’s behoorlijk hoger ligt dan in de natuur is waterverversing wel (erg) belangrijk. Bij temperaturen van 13 tot 16°C duurt de ontwikkeling van de 1 cm grote larve tot landfase ( ongeveer 4,5 a 5cm) doorgaans maximaal 2 maand, wat vergelijkbaar is met Ambystoma opacum. Voor een dergelijke snelle evolutie dient men echter wel frequent het water te verversen en voor een goede regelmatige voeding te zorgen.
Voor een goede groei van de dieren is het tevens belangrijker om regelmatig te voederen dan alle voeding ineens 1 maal per week aan te reiken. Vooral daphnia’s en later muggenlarven spelen een voorname rol. De grootte van de daphnia’s wordt afgespiegeld op de grootte van het oog van de larve. Om de goede grootte te verkrijgen, gebruik de artemia of daphnia zeefjes. Naar het einde van de metamorfose toe werden ook stukken regenworm gevoederd. De frequentie van de waterverversing dient opgedreven te worden naarmate de dieren groter worden, immers ze eten meer en zetten dus ook meer afvalstoffen af. Terwijl in het begin het water pas na 2 weken werd ververst, gebeurde dit naar het einde toe iedere 3 tot 4 dagen.
De eitjes werd in 2 verschillend ingerichte plastieken stapelbakjes opgekweekt : 1 met een grondlaag (A) en 1 zonder (B). Ietwat tegen mijn verwachtingen in bleken de larven in de kale bak B sneller te evolueren dan in bak A. De muggenlarven konden zich in bak B nl. niet ingraven, waardoor het voedsel veel langer aanwezig was. De constante aanwezigheid van voedsel betekent zeker een stevig pluspunt.
In de literatuur wordt regelmatig melding gemaakt van kannibalisme (Ambystoma macrodactylum columbianum). Zo noteerde Walls ea. (1993) dat zich tussen larven van deze ondersoort in het laboratorium 2 vormen ontwikkelden : normale larven met smalle kop en larven met een langere en bredere kop. Deze laatste bleken kannibalistische larven te zijn die ( bij voorkeur) op hun ‘kleinere’ soortgenoten aasden. Ook bij de Ambystoma tigrinum groep ontwikkelen zich dergelijke alternatieve larve-vormen, meerbepaald in de nominaatvorm , mavortium en nebulosum.

Nishihara rapporteert dat bij Hynobius retardatus een dergelijke kannibalistische vorm zich eerder onder hoge larven concentratie ontwikkeld (Nishihara, 1996). Andere auteurs die hierover ook rapporteren zijn : Powers 1907, Collins J.P. & Cheek J.E. 1983. Eigenaardig genoeg blijkt de predatie zich niet onder dieren van eenzelfde legsel voort te doen, men spreekt van ‘kinship recognition’ ( Pfennig D.W. & Collins J.P. 1993) m.a.w. broers en zussen herkennen elkaar als het ware. Bij mijn nakweken nam ik tot op heden geen cannibalisme waar, dit ofwel wegens de hoge voedingsfrequentie, ofwel vanwege het ‘kinship-recognition syndroom’.
Opkweek gemetamorfoseerde jongdieren
Eenmaal de dieren gemetamorfoseerd zijn werden ze ondergebracht in een kleine bak (20 op 30 cm). Een opkweekbakje voor net gemetamorfoseerde dieren kan een miniatuurversie zijn van de bak voor de adulte dieren. Hier moet echter nog meer dan bij de volwassen dieren toegezien te worden op een voldoende hoog aantal schuilplaatsen daar ze vooral bij het voeden vrij agressief tekeer gaan. Doorgaans wordt het bakje volgepropt met bouwstenen (met ventilatieopeningen) en pakken vochtig mos. Het bakje wordt hermetisch afgesloten zodat de (nodige) vochtigheid aanwezig blijft.
De jongen werden 2 tot 3 maal per week gevoederd. Telkenmale het bakje open ging kwamen de hapgrage mondjes boven. Niet zelden wordt zelfs voor dat voedsel in de bak aanwezig was naar elkaar gebeten, de bedoeling was duidelijk om zoveel mogelijk concurrentie op voorhand te verwijderen. Omdat het er soms ruw aan toe ging bracht ik de dieren onder in een bak met een drijfnatte bodem van sphagnum. Daarop werd vermolmd hout en mos geplaatst. Een dergelijk substraat is nl. vrij zuur en belet de ontwikkeling van bepaalde bacteriën.
Finaal slaagde ik erin om 48 dieren groot te brengen (50%). Deze werden verspreid over een aantal Franse, Nederlandse en Engelse salamanderliefhebbers. Een jaar later werd ik verwittigd dat de dieren zich reeds bij een Franse vriend hadden voortgeplant, ze waren toen amper meer dan 1,5 jaar oud. Ook ginds leverde de opkweek weinig problemen op. In 2000 mocht ik mijn eerste F-2 generatie opweken (25 stuks). Ook vanuit de Franse hoek blijkt dit gelukt te zijn.
Ziekte
Kort na de eerste nakweek ervaringen ontwikkelde zich in mijn adulte groep een schimmelinfectie op de staart van mijn dieren. Deze ziekte verspreidde zich (in de drijfnatte omgeving) razendsnel onder de andere dieren. Symptomen : op de staart ontwikkelde zicht een grote grijze schimmel, de staart viel af. Daarna ontwikkelden zich op het lichaam open wonden en de dieren stierven af.
Omdat ik op dat moment geen toegang had tot mensen met de nodige medische kennis probeerde ik –tevergeefs- om de dieren te redden door ze op verhakselde sphagnum grond te zetten. Met een gevoel van machteloosheid moest ik toezien hoe ik mijn volledige kweekgroep binnen een periode van 3 weken afstierf. Ik slaagde er gelukkig genoeg in om mijn nakweekdieren hiervoor te vrijwaren. Zoals hierboven vermeld zorgen deze ondertussen voor verdere nakweek, waardoor we deze dieren verder in de hobbykring kunnen behouden.
Literatuur
Anderson J.D., 1963, Reactions of the Western Mole to skin secretions of Ambystoma macrodactylum croceum, Herpetologica 19(4), 282/284
Anderson J.D. , 1968, A comparaison of the food habits of Ambystoma macrodactylum sigillatum, Ambystoma macrodactylum croceum and Ambystoma tigrinum californiense
Anderson J.D. 1972, Behavior of three subspecies of Ambystoma macrodactylum in a soil moisture gradient, Journal of Herpetology 6(3-4), 191/194
Collins, J.P., Cheek J.E., 1983, Effects of food and density on development of typical and cannibalistic salamander larvae in Ambystoma tigrinum nebulosum, Amer.Zool. 23(1) : 77-84.
Howard James H. & Wallace R.L, 1983, Critical Thermal Maxima in Populations of Ambystoma macrodactylum from different elevations, Copeia 400/402
Nishihara, A. 1996. High density induces a large head in larval Hynobius retardatus from a low density population. Japanese Journal of herpetology 16(4) : 134-136
Pfennig, D.W. en Collins J.P., 1993, Kinship affects morphogenesis in cannibalistic salamanders, Nature 362(6423) : 836-838.
Powers, J.H., 1907, Morphological variation and it’s causes in Amblystoma tigrinum, Stud.Univ.Nebraska 7(3) : 197-274.
Walls S.C, Beatty J.J., Tissot B.N., Hokit D.G., Blaustein A.R ., 1993, Morphological variation and cannibalism in a larval salamander (Ambystoma macrodactylum columbianum), Canad J. Zool (71), 1543/1551
Actueel

Nieuwe natuurpodcast: Schubben & Slijm
RAVON heeft een nieuwe podcast gelanceerd: Schubben & Slijm. In deze serie worden experts geïnterviewd over reptielen, amfibieën en vissen…

Veilige oversteek voor salamanders tijdens de voorjaarstrek
Elk voorjaar trekken niet alleen kikkers en padden, maar ook salamanders naar hun voortplantingswateren. Tijdens deze trek steken ze vaak…

Salamanders nog actief door zachte herfst
Het aanhoudend zachte weer zorgt ervoor dat veel salamanders hun winterslaap uitstellen. Vooral op vochtige avonden zijn ze nog te…